|
|
|
| CARVIEW |

 Kristin also produced a set of attractive anole (pus one boa) stickers, which she was handing out during her poster session.
Kristin also produced a set of attractive anole (pus one boa) stickers, which she was handing out during her poster session.


Traveling to SICB is always exciting, but like any trip through crowded airports, hotels, and convention centers, you’re more likely to get sick during your travel if you’re not careful. As we all know, getting a travel-cold (or worse) makes you feel terrible and certainly doesn’t make you want to run on a treadmill! The same is likely true in wild animals, including anoles. Mounting immune responses is energetically expensive, but so are other things that lizards have to do, like forage, escape predators, and process food. So, does an increasingly large immune challenge decrease a lizard’s ability to perform? Christine Rohlf, an undergraduate student in Jerry Husak’s lab at the University of St. Thomas wanted to find out in green anoles.
Christine designed a laboratory experiment to determine whether two types of immune challenge, alone and in combination, decreased bite-force performance, sprint speed, or endurance capacity compared to controls. Some lizards received two sequential injections of lipopolysaccharide (LPS), some received a skin wound with a biopsy punch, and some received both. LPS is a signal on gram-negative bacteria that, when injected, tricks the body into thinking it is infected with bacteria. So, you get an immune response, but you don’t actually get an infection.
Surprisingly, none of the immune challenges affected sprint speed or endurance compared to controls. Although the lizards were not calorie-restricted, they were on a modest diet, meaning that energy was limited, but clearly not enough to make a difference. Apparently these two immune challenges aren’t as costly as we thought. The only affect that Christine found was that the second LPS injection significantly decreased bite force. Because bite force is likely the least energetically expensive trait of those measured (imagine running until you’re exhausted versus biting into a hard piece of French bread), Christine suspects that the decrease in bite force was due to a lack of motivation while feeling sick. Future work with calorie-restricted lizards should tell us if mounting an immune challenge is a significant cost to anoles.
]]>
Amy Payne of Trinity University presents her research on tail autotomy in 7 lizard species.
One of the most interesting features of many lizards, including anoles, is that they can willingly, and actively, lose their tails to escape predators. While it might seem counterintuitive to lose a large body part, it’s better than being eaten! Despite the obvious benefit of surviving another day, there are some costs associated with tail autotomy.
Amy Payne, a student in Michele Johnson’s lab at Trinity University of San Antonio, wanted to know whether the frequency of tail loss across 7 species was associated with predatory and social use of the tail as well as energetic content of the tail. For those that anole-inclined (which is why you’re here), Amy included A. cristatellus and A. carolinensis. She caught and measured hundreds of lizards, and made behavioral observations on them as well. She was then able to quantify how many lizard of each species had a lost/regenerated tail, as well as what proportion of each tail was lost.
Surprisingly, frequency of tail loss was not associated with using the tail in a social or predatory context. However, using there was an association between these two functions of the tail: species that used their tail for predatory use more also used their tail in social contexts more. There was no relationship between the frequency of tail lost and the proportion of the tail that was lost on average across species. But she did find some really cool results when looking at energetic content of the tail. Amy found that there was a significant positive relationship between frequency of tail loss and tail energy content. That is, the more energy that lizards have in their tails, the more frequently individuals in that species will have a lost/regenerated tail. While this seems opposite to what one might casually predict, Amy hypothesizes that the predator-distraction to survive function of tail autotomy is more likely to succeed if the tail is larger and more beneficial to the predator. In other words, if a lizard has a scrawny tail and drops it off for a predator, it is more advantageous for the predator to ignore the low-cal tail and just eat the lizard. This would put selection on species with low-energy content tails to be more prudent about when they drop their tails. These really interesting results open up some exciting areas for future research on the costs and benefits of tail autotomy!
]]>In a recent study, as part of my PhD dissertation, I used a different approach to try to understand why most of these species are unable to colonize other islands. I used a recently developed conceptualization to link abundances and ecological niche requirements at coarse-grain scales, this approach has been developed in the lab of my advisor (see 4, 5, 6; but see 7, 8, 9 for discussions and counter-examples; this approach has been strongly debated in the literature in the last years).
We used ecological niche modeling -ENM- to predict species’ distributions across all Caribbean islands for each species with at least 10 occurrence records. We estimated the position of each pixel predicted as presence in the ecological space using Euclidean distances. In short, we characterize all pixels for a single species and calculated which of these were close to the niche centroid (which we assume as the best conditions for species presence) and which were close to the niche periphery (see Figure 1). We predicted that pixels predicted by ENM as presences within each native island will be more close to the niche centroid and those predicted as presences in other islands will be in the periphery of the niche.
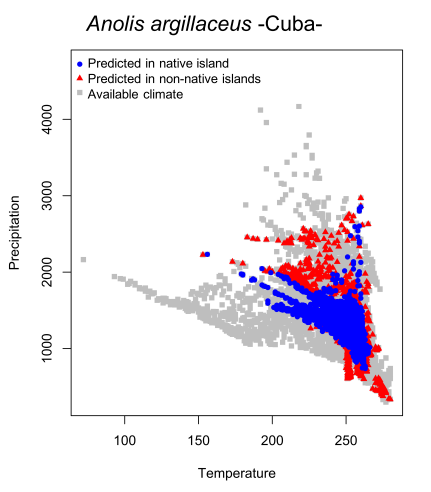
Figure 1. Available climatic space showing the position of each pixel predicted from ecological niche modeling across islands.
We found that many species follow the predicted pattern; in other words, we found that the “best” niche conditions are in the native islands regardless of climatic heterogeneity observed in each island and the “worst” niche conditions are outside native islands. We also used other metrics to corroborate our results. We interpreted these results as instances of recent climatic niche conservatism (within lineages) and therefore this operates as a constraint in the ability of each species to colonize other islands (i.e. due to the low suitable climatic conditions for successful population establishment). We only gathered data for 70 species and therefore it will be necessary more data and more studies (including physiological experiments) to corroborate our assertions.
Also, we examined the pattern of realised climatic niche shifts across the anole radiation and we found evidence of several instances of climatic niche convergence. We concluded that anoles evolved to occupy different portions of the climate space and in several cases evolved quickly to occupy some portions of this space (e.g., cold climatic conditions) and recently most of these species likely adapted very well to climatic conditions in its native islands.
The paper was published here:
https://link.springer.com/article/10.1007%2Fs11692-018-9455-x
]]>Thermal adaptation is the evolution of the ability to persist in novel thermal environments. Phenotypic characters that allow such adaptation, as well as the resulting shifts in the geographic distributions of species, are an emerging field of study in the midst of a changing global climate. Yet, the genomic basis of such phenotypic adaptation is less well understood, so recent efforts of evolutionary biologists are now aiming at one emerging question: Which genes determine thermal adaptation, and are these the same across different populations and species? Luckily, Anolis is yet again at the forefront of novel discoveries being made in this field (see Campbell-Staton et al., 2017).
Many studies have independently identified genes that are responding to changes in the thermal environment; be it through change of expression under an acute stress, or through changes in the DNA sequence as evolutionary response. In 2014, we gathered information on such thermal adaptation candidate genes from Drosophila to Homo sapiens from the literature. From the published evidence, we extracted a set of gene functions that potentially underlie climatic adaptation. We were able to match these with functions that we predicted from our observations of phenotypic thermal adaptation (Wollenberg Valero et al., 2014). Interestingly, the products of these genes (Proteins, RNAs) were found to be functionally related with each other thus forming gene networks within the cellular environment.
The Caribbean Anolis cybotes is widely distributed across Hispaniola, and thrives in hot, xeric environments just as well as in cooler and more humid montane environments. The rift valley of Lago Enriquillo heats up to 40.5 °C (104.9 °F), and a few instances of frost were reported at the highest peak (Pico Duarte at 3,098m elevation) – so population survival across these climatic extremes does not seem to be a trivial endeavor.

Anolis cybotes, female from Barahona, Dominican Republic
Populations of this species show pronounced differences between montane and lowland forms in morphology, physiology, behavior, and perch use (Wollenberg et al., 2013, Muñoz et al., 2014), which led us to expect that at least some of this variation should have a genetic basis. Thus, we set up to test whether Anolis cybotes displays any signatures of genomic adaptation to the diverse kinds of environments it inhabits, and whether any genes showing evidence for selection can also be subsumed under the candidate functions we defined previously.
We sampled tissue of these lizards from several high and low elevations (the specimens being the same as in Wollenberg et al., 2013), and looked for variation according to climatic differences via RAD sequencing and subsequent analysis with LFMM. RAD sequencing generates a reduced representation of the target genome, producing thousands of short sequences representing the distribution of the restriction enzyme’s cutting sites throughout the genome. Owing to this property, it cannot be expected that this type of data will necessarily contain “the total set of adaptation genes”; to this effect, detailed genome sequencing is required and such studies have been done in some model organisms (stickleback fish, beech mice, Drosophila, etc.). With our study design, however, we could trace signatures of selection as climate-related changes in the allelic frequencies in the fragments that were sequenced. We identified a total of 84 SNPs with statistical signatures of selection and 14 of these matched protein-coding genes on different chromosomes of the Anolis carolinensis genome (the best available reference). Not surprisingly, our data set and analysis did not “hit” any major known candidate genes for thermal adaptation, but we made another discovery. Most of the genes that we did identify as having adapted to the different climatic environments in Anolis cybotes populations perform the set of previously predicted gene functions that we had predicted in 2014 (Rodríguez et al., 2017).

The figure shows candidate gene functions for thermal adaptation that we predicted in 2014 and now verified in Anolis cybotes.
Moreover, the newly identified genes were also in close functional connection with each other, and with many of the previously predicted genes, formed a tightly knit functional network. Some forays into tissue expression databases further revealed that many of these genes are also expressed in brain, and during early development.
Due to the fact that we couldn’t scan the entire genome of Anolis cybotes (so far unknown), we may have missed part of the story. However, it is encouraging that at least one of the genes adapting to different climatic environments in the green anole, A. carolinensis, also has one of the functions we predicted (vasodilation, constriction and regulation of blood pressure, Campbell-Staton et al., 2016).
Phenotypic adaptation to climate can happen in several different ways and we are yet only scratching the surface of the genetic basis of this phenomenon. For reptiles, their thermoregulatory behavior, water balance in hot climates, freeze tolerance, and anti-oxidative strategies seem to be the most important eco-physiological challenges (Storey and Storey, 2017), and we deem it likely that changes in a multitude of genes are contributing to these adaptive responses. Our study provides evidence that climate adaptation on the genomic level is constrained to specific organismal functions and biochemical pathways, which may underpin the observed molecular and phenotypic differences. The definitive answer to the question on whether the same set of genes underlie climatic adaptation across populations and species, is yet to be found; but the study of the functional connections between genes can be very informative for this endeavor.
You can read our full paper here (open access)
]]>I can see that this behavior is rather common among the offsprings, and I partially film it in the following video. I didn’t see the same behavior in my others anoles species. Maybe only for trunk-ground species?
In which species did you see this behavior ?
In this case, the offspring begins to stiffen, twists and falls to the ground. The opens his mouth and stops breathing. The limbs are tense as a dead person. When young sent back safety (some seconds later) the individual’s “awakening”.
]]>I can see that this behavior is rather common among the offsprings, and I partially film it in this vidéo. I didn’t see the same behavior in my others anoles species, maybe only for trunk-ground species?
In which species did you see this behavior?
In this case, the offspring begins to stiffen, twists and falls to the ground. Then opens his mouth and stops breathing. The limbs are tense as a dead person. When young sent back safety (some seconds later) the individual’s “awakening”.
]]>I can see that this behavior is rather common among the offsprings, and I partially film it in this vidéo. In this case the offspring starts to stiffen, twists, and drops to the ground. Then opens his mouth and stops breathing. The limbs are tense as a dead individual.
A few seconds see minutes later, the individual of “awakening”.
It’s a recurrent problem in pet shop
In France a pet shop

 .
.
Thank you![]() for your interest
for your interest